MOVEMENT OF GASES AND WATER IN
VASCULAR PLANTS
The
Leaf
Although chloroplasts
are found in the cells of young stems and immature fruits, leaves are the real photosynthetic
factories of the plant.
A cross section through the
blade of a typical leaf reveals 4 distinct tissue layers.
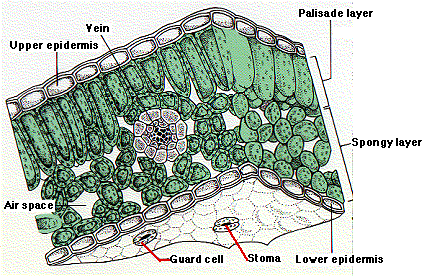 Upper
epidermis. This is a single
layer of cells containing few or no chloroplasts. The cells are quite
transparent and permit most of the light that strikes them to pass through
to the underlying cells. The upper surface is covered with a waxy,
waterproof cuticle, which serves to reduce water loss from the
leaf.
Upper
epidermis. This is a single
layer of cells containing few or no chloroplasts. The cells are quite
transparent and permit most of the light that strikes them to pass through
to the underlying cells. The upper surface is covered with a waxy,
waterproof cuticle, which serves to reduce water loss from the
leaf. - Palisade layer. This
consists of one or more layers of cylindrical cells oriented with their
long axis perpendicular to the plane of the leaf. The cells are filled
with chloroplasts (usually several dozen of them) and carry on most of the
photosynthesis
in the leaf.
- Spongy layer. Lying
beneath the palisade layer, its cells are irregular in shape and loosely
packed. Although they contain a few chloroplasts, their main function
seems to be the temporary storage of sugars
and amino
acids synthesized in the palisade layer. They also aid in the exchange
of gases between the leaf and the environment. During the day, these cells
give off oxygen and water vapor to the air spaces that surround
them. They also pick up carbon dioxide from the air spaces. The air spaces
are interconnected and eventually open to the outside through pores called
stomata (sing., stoma).
- Lower epidermis.
Typically. most of the stomata (thousands per square centimeter) are
located in the lower epidermis. Although most of the cells of the lower
epidermis resemble those of the upper epidermis, each stoma is flanked by
two sausage-shaped cells called guard cells. These differ from the
other cells of the lower epidermis not only in their shape but also in
having chloroplasts. The guard cells regulate the opening and closing of
the stomata. Thus they control the exchange of gases between the leaf and
the surrounding atmosphere.
|
|
Stomata under microscope taken from http://www.microscopy-uk.org.uk/mag/articles/stomata.html |
|
|
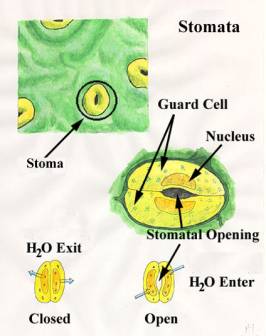
Diagram of stomata taken from http://www.puc.edu/Faculty/Gilbert_Muth/art0077.jpg |
 Leaf Veins
Leaf Veins
Not only must the cells of the
palisade and spongy layers be close to their air supply
- to secure CO2 and
- release O2
- (and the reverse in the dark)
but they must be close to a
leaf vein with its
The photo shows the network of leaf veins in
a maple leaf. Probably no cell in the spongy layer is more than two cells away
from a vein.
The xylem and phloem of veins is often
surrounded by layers of sclerenchyma
cells. These impart strength to the vein providing a stiff framework to support
the soft tissues of the leaf blade.
Gas Exchange in Plants
In order to carry on photosynthesis,
green plants need a supply of carbon dioxide and a means of disposing of
oxygen. In order to carry on cellular
respiration, plant cells need oxygen and a means of  disposing of carbon dioxide (just as animal
cells do).
disposing of carbon dioxide (just as animal
cells do).
Unlike animals, plants have
no specialized organs for gas exchange (with the few inevitable exceptions!).
The are several reasons they can get along without them:
- Each part of the plant takes care of its own gas exchange needs.
Although plants have an elaborate liquid transport system, it does not
participate in gas transport.
- Roots, stems, and leaves respire at rates much lower than are
characteristic of animals. Only during photosynthesis are large volumes of
gases exchanged and each leaf is well adapted to take care of its own
needs.
- The distance that gases must diffuse in even a large plant is not
great. Each living cell in the plant is located close to the surface.
While obvious for leaves, it is also true for stems. The only living
cells in the stem are organized in thin layers just beneath the bark. The
cells in the interior are dead and serve only to provide mechanical
support.
- Most of the living cells in a plant have at least part of their
surface exposed to air. The loose packing of parenchyma
cells in leaves, stems, and roots provides an interconnecting system of
air spaces. Gases diffuse through air several thousand times faster than
through water. Once oxygen and carbon dioxide reach the network of
intercellular air spaces (arrows), they diffuse rapidly through them.
- Oxygen and carbon dioxide also pass through the cell wall and
plasma membrane of the cell by diffusion. The diffusion of carbon dioxide
may be aided by aquaporin
channels inserted in the plasma membrane.
Leaves
The exchange of oxygen and carbon dioxide in
the leaf (as well as the loss of water vapor in transpiration)
occurs through pores called stomata (singular = stoma).
Normally stomata open when
the light strikes the leaf in the morning and close during the night.
The immediate cause is a change in the turgor
of the guard cells. The inner wall of each guard cell is thick and
elastic. When turgor develops within the two guard cells flanking each stoma,
the thin outer walls bulge out and force the inner walls into a crescent shape.
This opens the stoma. When the guard cells lose turgor, the elastic inner walls
regain their original shape and the stoma closes.
|
Time |
Osmotic Pressure, lb/in2 |
|
7 A.M. |
212 |
|
11 A.M. |
456 |
|
5 P.M. |
272 |
|
12 midnight |
191 |
The table shows the osmotic pressure
measured at different times of day in typical guard cells. The osmotic pressure
within the other cells of the lower epidermis remained constant at 150 lb/in2.
When the osmotic pressure of the guard cells became greater than that of the
surrounding cells, the stomata opened. In the evening, when the osmotic
pressure of the guard cells dropped to nearly that of the surrounding cells,
the stomata closed.
Opening stomata
The increase in osmotic
pressure in the guard cells is caused by an uptake of potassium ions (K+).
The concentration of K+ in open
guard cells far exceeds that
in the surrounding cells. This is how it accumulates:
- Blue light is absorbed by phototropin
which activates
- a proton
pump (an H+-ATPase) in the plasma membrane of the guard
cell.
- ATP, generated by the light
reactions of photosynthesis, drives the pump.
- As protons (H+) are pumped out of the cell, its interior
becomes increasingly negative.
- This attracts additional potassium ions into the cell, raising its
osmotic pressure.
Closing stomata
Although open stomata are essential for
photosynthesis, they also expose the plant to the risk of losing water through transpiration.
Some 90% of the water taken up by a plant is lost in transpiration.
Abscisic
acid (ABA) is the hormone that triggers closing of the stomata when soil
water is insufficient to keep up with transpiration (which often occurs around
mid-day).
The mechanism:
- ABA binds to receptors at the surface of the plasma membrane of the
guard cells.
- The receptors activate several interconecting pathways which
converge to produce
- The increased Ca2+ in the cytosol blocks the uptake of K+
into the guard cell while
- the increased pH stimulates the loss of Cl- and organic
ions (e.g., malate2-) from the cell.
- The loss of these solutes in the cytosol reduces the osmotic
pressure of the cell and thus turgor.
- The stomata close.
Density of stomata
The density of stomata on a
leaf varies with such factors as:
- the temperature, humidity, and light intensity
around the plant;
- and also, as it turns out, the concentration of carbon dioxide
in the air around the leaves. The relationship is inverse; that is,
as CO2 goes up, the number of stomata goes down, and vice
versa. Some evidence:
- Plants grown in an artificial atmosphere with a
high level of CO2 have fewer stomata than normal.
- Herbarium specimens reveal that the number of
stomata in a given species has been declining over the last 200 years —
the time of the industrial revolution and rising levels of CO2
in the atmosphere.
These data can be quantified by determining
the stomatal index: the ratio of the number of stomata in a given area
divided by the total number of stomata and other epidermal cells in that same
area.
How does the plant determine how many
stomata to produce?
It turns out that the mature leaves on the
plant detect the conditions around them and send a signal (its nature still
unknown) that adjusts the number of stomata that will form on the developing
leaves.
Two experiments (reported by Lake et al., in
Nature, 411:154, 10 May 2001):
- When the mature leaves of the plant (Arabidopsis)
are encased in glass tubes filled with high levels (720 ppm)
of CO2, the developing leaves have fewer stomata than normal
even though they are growing in normal air (360 ppm).
- Conversely, when the mature leaves are given normal air (360 ppm CO2)
while the shoot is exposed to high CO2 (720 ppm), the new
leaves develop with the normal stomatal index.
Stomata reveal past carbon dioxide levels
Because CO2
levels and stomatal index are inversely related, could fossil leaves tell us
about past levels of CO2 in the atmosphere? Yes. As reported by
Gregory Retallack (in Nature, 411:287, 17 May 2001), his study of
the fossil leaves of the ginkgo
and its relatives shows:
- their stomatal indices were high
- late in the Permian
period (275–290 million years ago) and again
- in the Pleistocene
epoch (1–8 million years ago).
Both these periods are known
from geological evidence to have been times of
- low
levels of atmospheric carbon dioxide and
- ice ages (with glaciers).
- Conversely, stomatal indices were low during the Cretaceous
period, a time of high CO2 levels and warm climate.
These studies also lend
support to the importance of carbon dioxide as a greenhouse gas playing an
important role in global
warming.
 Roots and Stems
Roots and Stems
Woody stems and mature roots are sheathed in
layers of dead cork cells impregnated with suberin — a waxy, waterproof
(and airproof) substance. So cork is as impervious to oxygen and carbon dioxide
as it is to water.
However, the cork of both
mature roots and woody stems is perforated by nonsuberized pores called lenticels.
These enable oxygen to reach the intercellular spaces of the interior tissues
and carbon dioxide to be released to the atmosphere.
The photo shows the lenticels in the bark of
a young stem.
In many annual plants, the stems are green
and almost as important for photosynthesis as the leaves. These stems use
stomata rather than lenticels for gas exchange.
Transpiration
Transpiration is the evaporation of water from
plants. It occurs chiefly at the leaves while their stomata
are open for the passage of CO2 and O2 during photosynthesis.
But air that is not fully saturated with
water vapor (100% relative humidity) will dry the surfaces of cells with which
it comes in contact. So the photosynthesizing leaf loses substantial amount of
water by evaporation. This transpired water must be replaced by the transport
of more water from the soil to the leaves through the xylem
of the roots and stem.
Importance
Transpiration is not simply
a hazard of plant life. It is the "engine" that pulls water up from
the roots to:
- supply photosynthesis (1%-2% of the total)
- bring minerals from the roots for biosynthesis within the leaf
- cool the leaf
Environmental
factors that affect the rate of transpiration
1. Light
Plants transpire more rapidly in the light than in the dark. This is largely
because light stimulates the opening of the stomata (mechanism).
Light also speeds up transpiration by warming the leaf.
2. Temperature
Plants transpire more rapidly at higher temperatures because water evaporates
more rapidly as the temperature rises. At 30°C, a leaf may transpire three
times as fast as it does at 20°C.
3. Humidity
The rate of diffusion
of any substance increases as the difference in concentration of the substances
in the two regions increases. When the surrounding air is dry, diffusion of
water out of the leaf goes on more rapidly.
4. Wind
When there is no breeze, the air surrounding a leaf becomes increasingly humid
thus reducing the rate of transpiration. When a breeze is present, the humid
air is carried away and replaced by drier air.
5. Soil water
A plant cannot continue to transpire rapidly if its water loss is not made up
by replacement from the soil. When absorption of water by the roots fails to
keep up with the rate of transpiration, loss of turgor
occurs, and the stomata close. This immediately reduces the rate of
transpiration (as well as of photosynthesis). If the loss of turgor extends to
the rest of the leaf and stem, the plant wilts.
The volume of water lost in transpiration
can be very high. It has been estimated that over the growing season, one acre
of corn plants may transpire 400,000 gallons of water. As liquid water, this
would cover the field with a lake 15 inches deep. An acre of forest probably
does even better.
Transport
of Water and Minerals in Plants
Most plants secure the water and minerals
they need from their roots.
The path taken is: soil -> roots ->
stems -> leaves
The minerals (e.g., K+, Ca2+)
travel dissolved in the water (often accompanied by various organic molecules
supplied by root cells).
Less than 1% of the water reaching the
leaves is used in photosynthesis and plant growth. Most of it is lost in
transpiration.
However, transpiration does serve two useful
functions:
- It provides the force for lifting the water up the stems.
- It cools the leaves.
Water and minerals enter the root by
separate paths which eventually converge in the stele.
The Pathway of Water
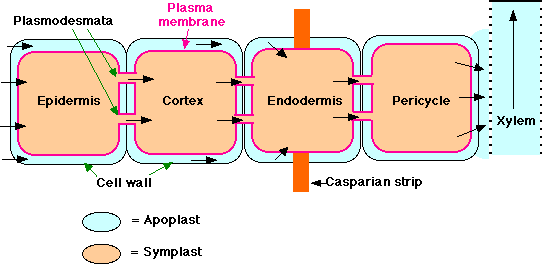
Soil water enters the root through its
epidermis. It appears that water then travels in both
- the cytoplasm of root cells — called the symplast
— that is, it crosses the plasma membrane and then passes from cell to
cell through plasmodesmata.
- in the nonliving parts of the root — called the apoplast —
that is, in the spaces between the cells and in the cells walls
themselves. This water has not crossed a plasma membrane.
However, the inner boundary of the cortex,
the endodermis, is impervious to water because of a band of suberized
matrix called the casparian strip. Therefore, to enter the stele,
apoplastic water must enter the symplasm of the endodermal cells. From here it
can pass by plasmodesmata into the cells of the stele.
Once inside the stele, water is again free
to move between cells as well as through them. In young roots, water enters
directly into the xylem vessels and/or tracheids. These are nonliving
conduits so are part of the apoplast.
Once in the xylem, water with the minerals that
have been deposited in it (as well as occasional organic molecules supplied by
the root tissue) move up in the vessels and tracheids.
At any level, the water can leave the xylem
and pass laterally to supply the needs of other tissues.
At the leaves, the xylem passes into the
petiole and then into the veins of the leaf. Water leaves the finest veins and
enters the cells of the spongy
and palisade layers. Here some of the water may be used in metabolism, but
most is lost in transpiration.
The Pathway of Minerals
Minerals enter the root by active
transport into the symplast of epidermal cells and move toward and into the
stele through the plasmodesmata connecting the cells.
They enter the water in the xylem from the
cells of the pericycle (as well as of parenchyma cells surrounding the xylem)
through specialized transmembrane channels.
What Forces Water Through the
Xylem?
Observations
- The mechanism is based on purely physical forces because the xylem
vessels and tracheids are lifeless.
- Roots are not needed. This was demonstrated over a century ago by a
German botanist who sawed down a 70-ft oak tree and placed the base of the
trunk in a barrel of picric acid solution. The solution was drawn up the
trunk, killing nearby tissues as it went.
- However, leaves are needed. When the acid reached the leaves and
killed them, the upward movement of water ceased.
- Removing a band of bark
from around the trunk — a process called girdling — does not
interrupt the upward flow of water. Girdling removes only the phloem, not
the xylem, and so the foliage does not wilt. (In due course, however, the
roots — and thus the entire plant — will die because the roots cannot
receive any of the food manufactured by the leaves.)
In 1895, the Irish plant physiologists H. H.
Dixon and J. Joly proposed that water is pulled up the plant by tension
(negative pressure) from above.
As we have seen, water is continually being
lost from leaves by transpiration. Dixon and Joly believed that the loss of
water in the leaves exerts a pull on the water in the xylem ducts and draws
more water into the leaf.
But even the best vacuum pump can pull water
up to a height of only 34 ft or so. This is because a column of water that high
exerts a pressure (~15 lb/in2) just counterbalanced by the pressure
of the atmosphere. How can water be drawn to the top of a sequoia (the tallest
is 370 feet high)? Taking all factors into account, a pull of at least 270
lb/in2 is probably needed.
The answer to the dilemma lies the cohesion
of water molecules; that is the property of water molecules to cling to each
through the hydrogen bonds they form.
When water is confined to tubes of very
small bore, the force of cohesion between water molecules imparts great
strength to the column of water. Tensions as great as 3000 lb/in2
are needed to break the column, about the value needed to break steel wires of
the same diameter. In a sense, the cohesion of water molecules gives them the
physical properties of solid wires.
Because of the critical role of cohesion,
the transpiration-pull theory is also called the cohesion theory.
Some support for the theory
- If sap in the xylem is under tension, we would expect the column to
snap apart if air is introduced into the xylem vessel by puncturing it.
This is the case.
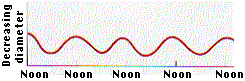
- If the water in all the xylem ducts is under tension, there should
be a resulting inward pull (because of adhesion) on the walls of the
ducts. This inward pull in the band of sapwood in an actively transpiring
tree should, in turn, cause a decrease in the diameter of the
trunk.
The graph shows the
results of obtained by D. T. MacDougall when he made continuous measurements of
the diameter of a Monterey pine. The diameter fluctuated on a daily basis
reaching its minimum when the rate of transpiration reached its maximum
(around noon)
- The rattan vine may climb as high as 150 ft on the trees of the
tropical rain forest in northeastern Australia to get its foliage into the
sun. When the base of a vine is severed while immersed in a basin of
water, water continues to be taken up. A vine less than 1 inch in diameter
will "drink" water indefinitely at a rate of up to 12 ml/minute.
If forced to take
water from a sealed container, the vine does so without any decrease in rate,
even though the resulting vacuum becomes so great that the remaining water
begins to boil spontaneously. (The boiling temperature of water decreases as
the air pressure over the water decreases, which is why it takes longer to boil
an egg in Denver than in New Orleans.)
- Transpiration-pull enables some trees and shrubs to live in
seawater. Seawater is markedly hypertonic
to the cytoplasm in the roots of the coastal mangrove, and we might expect
water to leave the cells resulting in a loss in turgor
and wilting. In fact, the remarkably high tensions (on the order of
500–800 lb/in2) in the xylem can pull water into the plant
against this osmotic
gradient. So mangroves literally desalt seawater to meet their needs.
Problems with the theory
When water is placed under a high vacuum,
any dissolved gases come out of solution as bubbles (as we saw above with the
rattan vine). This is called cavitation. Any impurities in the water
enhance the process. So measurements showing the high tensile strength of water
in capillaries require water of high purity — not the case for sap in the
xylem.
So might cavitation break the column of
water in the xylem and thus interrupt its flow? Probably not so long as the
tension does not greatly exceed 270 lb/in2.
By spinning branches in a centrifuge, it has
been shown that water in the xylem avoids cavitation at negative pressures
exceeding 225 lb/in2. And the fact that sequoias can successfully
lift water 358 ft (109 m) — which would require a tension of 270 lb/in2
— indicates that cavitation is avoided even at that value.
However, such heights may be
approaching the limit for xylem transport. (The tallest tree ever measured, a
Douglas fir, was 413 ft. high.) Measurements close to the top of the tallest
living sequoia (370 ft high) show that the high tensions needed to get water up
there have resulted in:
- smaller stomatal
openings, causing
- lower concentrations of CO2 in the needles, causing
- reduced photosynthesis, causing
- reduced growth (smaller cells and much smaller needles).
(See Koch, G. W. et al.,
Nature, 22 April 2004.)
Root Pressure
When a tomato plant is
carefully severed close to the base of the stem, sap oozes from the stump. The
fluid comes out under pressure which is called root pressure.
Root pressure is created by the osmotic
pressure of xylem sap which is, in turn, created by dissolved
- minerals and
- sugars
that have been actively
transported into the apoplast of the stele.
One important example is the sugar maple
when, in very early spring, it hydrolyzes
the starches stored in its roots into sugar. This causes water to pass by
osmosis through the endodermis and into the xylem ducts. The continuous inflow
forces the sap up the ducts.
Although root pressure plays a role in the
transport of water in the xylem in some plants and in some seasons, it does not
account for most water transport.
- Few plants develop root pressures greater than 30 lb/in2,
and some develop no root pressure at all.
- The volume of fluid transported by root pressure is not enough to
account for the measured movement of water in the xylem of most trees and
vines.
- Those plants with a reasonably good flow of sap are apt to have the
lowest root pressures and vice versa.
- The highest root pressures occur in the spring when the sap is
strongly hypertonic
to soil water, but the rate of transpiration is low. In summer, when
transpiration is high and water is moving rapidly through the xylem, often
no root pressure can be detected.
So although root pressure may play a
significant role in water transport in certain species (e.g., the coconut palm)
or at certain times, most plants meet their needs by transpiration-pull.
Adapted from Kimball's Biology Pages: http://users.rcn.com/jkimball.ma.ultranet/BiologyPages